Introduction
The plasma membrane (also called the cell membrane or plasmalemma) is a thin, flexible, selectively permeable membrane that surrounds the cytoplasm of every living cell. It forms the boundary between the internal environment of the cell and the external environment, regulating the movement of substances into and out of the cell while maintaining cellular integrity.
Structurally, the plasma membrane consists primarily of a phospholipid bilayer embedded with proteins, cholesterol (in animal cells), and carbohydrates, following the Fluid Mosaic Model proposed by Singer and Nicolson (1972).
Importance of the Plasma Membrane
The plasma membrane is essential for the survival and proper functioning of all living cells. It maintains structural integrity and shapes the cell while completely separating the internal environment from the outside. Acting as a careful checkpoint, it selectively controls the entry of important nutrients, water, and ions. It also allows metabolic wastes to exit, which helps keep the cell in balance. Crucially, by regulating this continuous molecular traffic and balancing internal conditions, the membrane acts as the primary driver of cellular homeostasis.
The membrane does more than just transport substances. It supports cell recognition, immune responses, and complex communication between cells through its surface carbohydrates and specialized receptors. Additionally, it provides important physical support for both the internal cytoskeleton and the external extracellular matrix. It also plays a role in bulk transport processes like endocytosis and exocytosis. Finally, it maintains the precise electrochemical gradients needed for nerve impulse transmission and muscle contractions.
Singer and Nicolson's Fluid Mosaic Model (1972)
Proposed by S. Jonathan Singer and Garth Nicolson
The plasma membrane is described as a fluid phospholipid bilayer with proteins embedded within it.
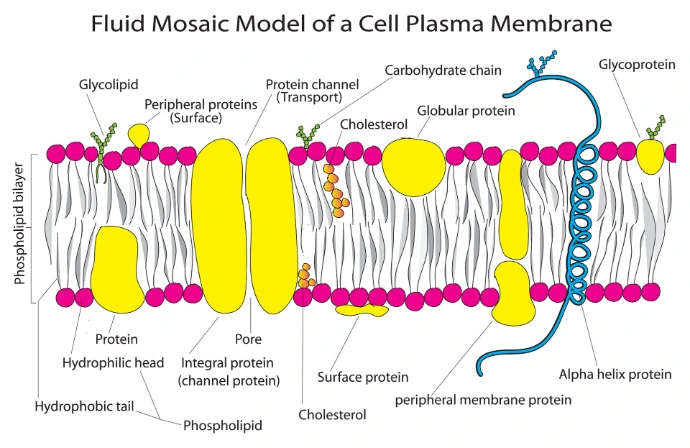
Arrangement in the Plasma Membrane
In an aqueous environment, phospholipids spontaneously arrange themselves into a bilayer.
• Hydrophilic heads face the extracellular fluid and cytoplasm.
• Hydrophobic tails face inward toward one another, forming a water-repellent interior.
This arrangement creates a stable and selectively permeable membrane.
Main Features:
• Phospholipids form a bilayer.
• Membrane proteins float within the lipid bilayer like a mosaic.
• Lipids and many proteins move laterally, making the membrane dynamic.
• Cholesterol regulates membrane fluidity (especially in animal cells).
• Carbohydrates are attached to proteins and lipids on the outer surface, forming the glycocalyx.
• The membrane is selectively permeable, allowing controlled movement of substances.
Chemical Composition of the Plasma Membrane–Lipids
The main types of membrane lipids are phospholipids, glycolipids, sphingolipids, cholesterol (in animals), plant sterols, and hopanoids (in bacteria).
Phospholipids
Phospholipids are the most abundant membrane lipids and form the structural backbone of the plasma membrane. They are amphipathic molecules, meaning they possess both a hydrophilic (water-attracting) region and a hydrophobic (water-repelling) region.
Each phospholipid molecule consists of:
• Glycerol backbone
• Two fatty acid tails (hydrophobic/non-polar)
• One phosphate group attached to a polar alcohol such as choline, ethanolamine, serine or inositol (hydrophilic/polar head)
Thus, each molecule has:
• Polar (hydrophilic) head
• Non-polar (hydrophobic) tails
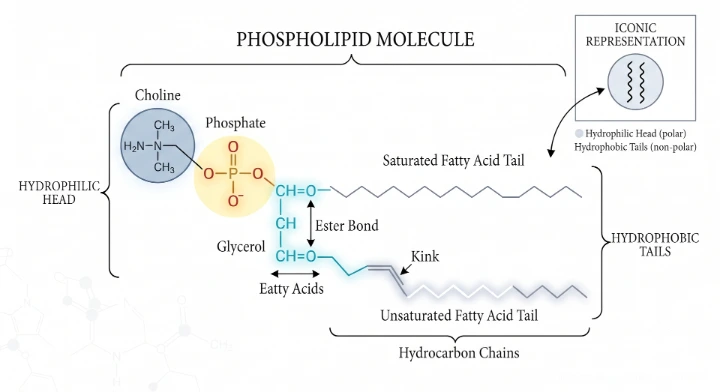
Types of Phospholipids
• Common phospholipids include:
• Phosphatidylcholine (Lecithin)
• Phosphatidylethanolamine
• Phosphatidylserine
• Phosphatidylinositol
• Cardiolipin (mainly in mitochondria)
Glycolipids
Glycolipids are specialized membrane lipids in which one or more carbohydrate chains are covalently attached to lipid molecules. They exhibit strict structural asymmetry, being present only in the outer leaflet of the plasma membrane. From this position, their carbohydrate portions project outward from the cell surface into the extracellular fluid, serving as a major component of the glycocalyx (cell coat).
Structure
To understand a glycolipid, it must be viewed as an amphipathic molecule divided into two distinct functional regions:
The Carbohydrate Component (Hydrophilic Head): This polar region consists of one or more sugar residues attached to the membrane surface. These can range from simple monosaccharides like glucose or galactose to complex, highly branched oligosaccharides.
The Lipid Component (Hydrophobic Tail): The carbohydrate head is anchored to the bilayer by being covalently bonded to one of two structural lipid bases:
•Glycerol-based (Glyceroglycolipids): The sugar chain attaches to a glycerol molecule bound to two fatty acid tails (predominant in plant and bacterial membranes).
•Sphingosine-based (Glycosphingolipids): The sugar chain attaches to a sphingosine backbone bound to a fatty acid tail—a combination collectively known as a ceramide unit (predominant in animal cell membranes, especially nervous tissue).
The Covalent Linkage: The chemical bond connecting the sugar residue directly to the polar head of the lipid backbone is a specific glycosidic bond (specifically an O-glycosidic bond).
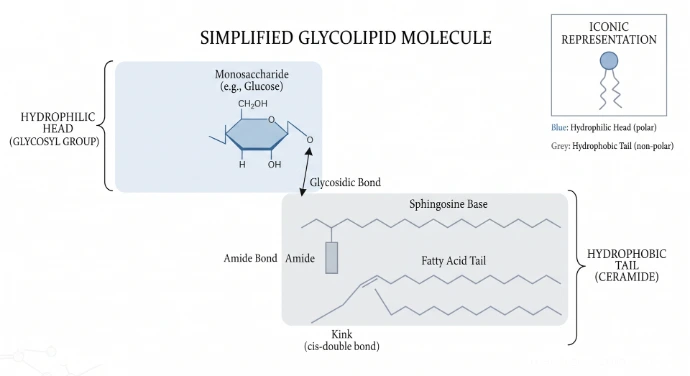
Function
• Cell recognition.
• Cell-to-cell communication.
• Cell adhesion.
• Immune recognition.
• Protection of the cell surface.
• Stabilization of the membrane.
Sphingolipids
Sphingolipids are a complex class of structural membrane lipids characterized by the presence of a sphingosine backbone instead of the glycerol framework found in standard phospholipids. They are ubiquitous components of eukaryotic plasma membranes but are uniquely concentrated within the nervous system—particularly within the myelin sheath that wraps and electrically insulates nerve fibers. Because of this specialized distribution, sphingolipids are absolutely critical for structural neural insulation, cellular signaling, and the healthy physiological functioning of the brain.
Structure
A sphingolipid molecule is structurally organized into three core chemical building blocks:
• The Sphingosine Backbone: A long-chain, structural amino alcohol that serves as the foundation of the molecule.
• A Single Fatty Acid Tail: A hydrophobic hydrocarbon chain attached directly to the amino group (-NH2) of the sphingosine backbone via an amide bond. The combination of this single fatty acid chain and the sphingosine backbone form a hydrophobic core structure known as a ceramide.
• A Polar Head Group: A hydrophilic functional group esterified or linked to the terminal hydroxyl (-OH) group of the sphingosine base. The specific identity of this head group dictates the classification of the sphingolipid molecule.
Major types include:
• Sphingomyelin
• Cerebrosides
• Gangliosides
Function
• Increase membrane stability.
• Participate in cell signaling.
• Form lipid rafts with cholesterol.
• Insulate nerve fibers.
• Facilitate cell recognition and communication.

Cholesterol
Cholesterol is the principal sterol present in animal cell membranes, serving as a vital architectural component that constitutes up to 30–50% of the total membrane lipid fraction in some animal cells. It exhibits highly kingdom-specific distribution: it is completely absent from most bacterial membranes (which utilize hopanoids instead) and occurs in only trace amounts in plant cell membranes (which rely on phytosterols)
Structure
The cholesterol molecule possesses a unique, rigid architecture that dictates exactly how it nests inside the fluid membrane:
• The Four Fused Hydrocarbon Rings: A bulky, planar, and rigid steroid ring system that forms the core of the molecule.
• A Short Hydrocarbon Tail: A flexible, non-polar tail extending from one end of the ring system.
• A Single Hydroxyl (-OH) Group: A small polar head group located at the opposite end of the molecule.
Because of this specific structural layout, cholesterol aligns itself asymmetrically and vertically within the lipid bilayer. The small, hydrophilic hydroxyl (-OH) group forms hydrogen bonds directly with the polar heads of surrounding phospholipids. Concurrently, the flat, rigid steroid rings and the flexible hydrocarbon tail plunge into the hydrophobic core, inserting themselves parallel to the fatty acid tails.
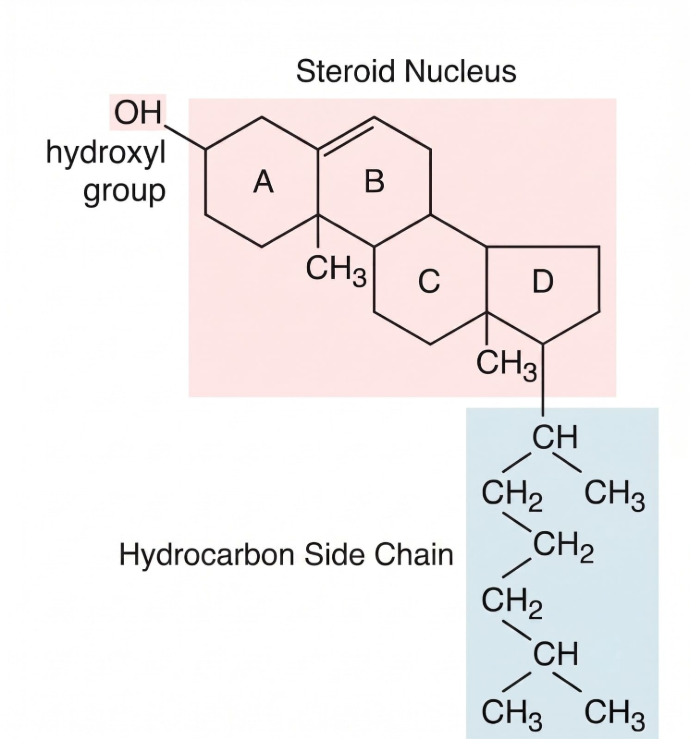
Function
Regulates membrane fluidity
• Prevents excessive rigidity at low temperatures.
• Prevents excessive fluidity at high temperatures.
Stabilizes the membrane
Provides mechanical strength and reduces membrane deformation.
Reduces membrane permeability
Decreases leakage of small water-soluble molecules.
Organizes lipid rafts
Works with sphingolipids to form specialized membrane domains involved in signaling.
Phytosterols: Sterols in Plants
While animal cells rely entirely on cholesterol, plant cell membranes utilize a diverse group of steroid alcohols known as phytosterols (plant sterols) as their primary architectural sterols. They are synthesized within the endoplasmic reticulum and populate the plasma membrane to regulate structural mechanics.
Common Plant Sterols
• β-Sitosterol
• Stigmasterol
• Campesterol
Structural Comparison with Cholesterol
Phytosterols are structural analogs of cholesterol, sharing the exact same hydrophobic four-ring core and hydrophilic hydroxyl (-OH) group. However, they differ slightly in chemical structure due to additional alkyl groups (such as methyl or ethyl groups) or unique double bonds located on their hydrocarbon side chains. Despite these minor modifications, they insert themselves into the plant phospholipid bilayer in the exact same manner as cholesterol does in animal cells.
Function
• Maintain membrane fluidity.
• Stabilize the phospholipid bilayer.
• Reduce membrane permeability.
• Support growth and development.
• Help plants tolerate temperature stress.
Hopanoids in Bacteria
Hopanoids are a highly specialized class of pentacyclic triterpenoid lipids synthesized by a wide variety of prokaryotes. Because the vast majority of bacteria lack the biochemical pathways required to produce eukaryotic sterols like cholesterol, they rely on hopanoids as functional architectural equivalents to reinforce and modulate their plasma membranes.
Structure
Unlike the four fused hydrocarbon rings seen in cholesterol, the chemical backbone of a hopanoid consists of five fused hydrocarbon rings (a pentacyclic ring system). This rigid, bulky core allows the molecule to align perfectly within the bacterial lipid bilayer. Just like cholesterol, hopanoids insert themselves parallel to the fatty acid chains of phospholipids, acting as structural anchors that tightly control the physical dynamics of the membrane matrix.
Microbial Distribution
• Widespread in Bacteria: Hopanoids are highly abundant in structurally diverse bacterial groups, particularly cyanobacteria, nitrogen-fixing bacteria (such as Bradyrhizobium), and various groups of soil bacteria that frequently encounter severe environmental fluctuations.
• Absent in Archaea: Archaea do not contain hopanoids; instead, they adapt to extreme environments using entirely distinct, highly stable ether-linked membrane lipids characterized by branched isoprenoid chains.
Function
• Increase membrane stability.
• Regulate membrane fluidity.
• Reduce membrane permeability.
• Protect bacteria from environmental stress.
• Improve membrane rigidity under harsh conditions.
Membrane Proteins
Membrane proteins are proteins associated with the plasma membrane that perform numerous structural and functional roles. Proteins constitute approximately 50–60% of the plasma membrane by mass, although this proportion varies among different cell types.
Membrane proteins may be:
• Embedded within the lipid bilayer
• Attached to its surface
• Anchored by lipid molecules
Classification of Membrane Proteins
• Integral Proteins (Intrinsic Proteins)
• Peripheral Proteins (Extrinsic Proteins)
• Lipid-Anchored Proteins

Structural Features of Integral Membrane Proteins
Integral membrane proteins possess specialized structural features that enable them to interact with, embed within, and often span the phospholipid bilayer. These features are primarily determined by the distribution of hydrophobic and hydrophilic amino acid residues within the protein. The major structural features include hydrophobic regions and transmembrane domains.
Hydrophobic Regions
Hydrophobic regions are segments of an integral membrane protein that are rich in nonpolar amino acids. These regions interact favorably with the hydrophobic fatty acid tails of phospholipids, allowing the protein to associate with the lipid bilayer.
Characteristics
• Rich in hydrophobic (nonpolar) amino acids.
• Interact with the hydrophobic core of the membrane.
• Help anchor and stabilize proteins within the lipid bilayer.
• May be partially embedded within the membrane or form part of a transmembrane domain.
Functions
• Anchor proteins within the lipid bilayer.
• Stabilize protein–membrane interactions.
• Facilitate interactions with membrane lipids.
• Form the hydrophobic core of transmembrane domains
Transmembrane Domains
A transmembrane domain is a specialized hydrophobic region of an integral membrane protein that extends completely across the phospholipid bilayer, connecting the extracellular and intracellular environments.
Characteristics
• Span the entire phospholipid bilayer.
• Typically consist of 20–25 hydrophobic amino acid residues.
• Form stable interactions with membrane lipids.
• Can occur as a single transmembrane segment or as multiple transmembrane segments.
Types of Transmembrane Domains
Single-pass Transmembrane Proteins
These proteins possess a single transmembrane domain and cross the membrane only once.
Multi-pass Transmembrane Proteins
These proteins contain multiple transmembrane domains, allowing them to cross the membrane several times.
Structural Architectures of Transmembrane Domains
Transmembrane domains are primarily organized into two structural architectures
• α-Helical Transmembrane Domains
• β-Barrel Transmembrane Domains
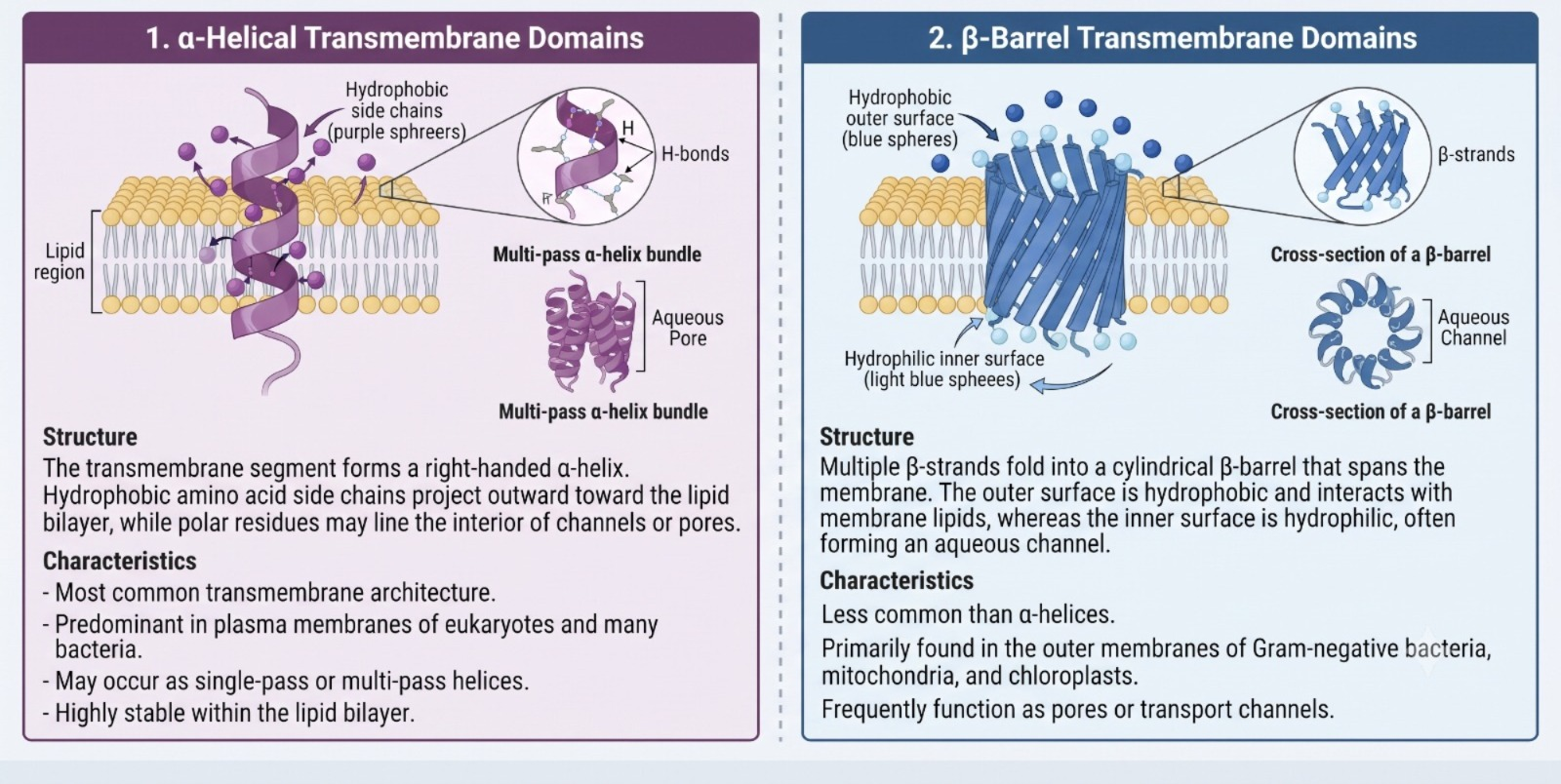
Conclusion
The plasma membrane serves as a highly sophisticated, dynamic molecular gatekeeper essential for cellular survival and homeostasis. Following the Fluid Mosaic Model, it balances structural flexibility with robust barrier integrity. The foundational phospholipid bilayer is reinforced by kingdom-specific architectural modulators—cholesterol in animals, phytosterols in plants, and hopanoids in bacteria—which actively buffer membrane fluidity against environmental stresses. Concurrently, specialized surface glycolipids and sphingolipids build the protective glycocalyx coat, orchestrating crucial cellular operations including immune surveillance, tissue adhesion, and neural insulation.
Embedded directly within this fluid lipid matrix, membrane proteins function as the primary functional machinery of the cell border. Spanning the bilayer via single-pass or multi-pass transmembrane domains, these integral and peripheral structures execute complex physiological tasks. They maintain precise electrochemical gradients, anchor the internal cytoskeleton, and serve as the exact channels, pumps, and receptors required for selective transport and intercellular communication, altogether forming the structural foundation of life.